Fra Jerry Coyne, "Evolution-of-the-Gaps" og andre feilslutninger
Jonathan McLatchie 5. desember 2012 Evolusjon, intelligent design. Oversatt herfra
 Bilde 1. Sekvensiell DNA-kode kan ikke oppstå naturlig
Bilde 1. Sekvensiell DNA-kode kan ikke oppstå naturlig
Fortsetter min anmeldelse av Jerry Coynes Hvorfor evolusjon er sann.
Kapittel 3: Rester - Spor, embryoer og dårlig design
I kapittel 3 i boken sin vender Coyne oppmerksomheten mot argumentet fra suboptimal design. Mens feilslutningen "gud-i-hullene" sier at "evolusjon ikke kan forklare dette; derfor må Gud ha gjort det", begår Coynes argument i dette kapittelet en omvendt feilslutning "evolusjon-av-hullene": "Gud ville ikke ha gjort det på den måten; derfor må evolusjonen ha gjort det." Det er merkelig at denne dikotome tankegangen er nettopp det darwinister som Coyne så ofte liker å anklage ID-tilhengere for. I likhet med argumentene om "gud-i-hullene", må argumentet om "evolusjon-i-hullene" vike med fremskritt innen vitenskapelig kunnskap, ettersom biologer avdekker viktige årsaker til måten disse funksjonene har blitt designet på. Videre er en vanlig kritikk av ID at den er uvitenskapelig fordi den ikke er falsifiserbar. Men i dette kapittelet hevder Coyne å ha falsifisert designhypotesen. Man kan ikke ha det begge veier - enten er ID falsifiserbar eller ikke. Ved å tilby en kritikk av design som en hypotese, antyder Coyne at ID er falsifiserbar, og derfor at den er vitenskapelig etter det kriteriet.
Rester
 Det første eksemplet på et rudimentært trekk som er sitert her, er vingene til strutser og andre fugler som ikke kan fly, som strutsfugler. Få vil tvile på at en struts' forfedre var flygende fugler. Dette er imidlertid ikke et veldig sterkt bevis, hvis Coyne ønsker å overbevise skeptikere om effektiviteten til moderne evolusjonsteori - rudimentære strukturer av denne typen dokumenterer i beste fall tap av funksjon. Det som må demonstreres er at moderne evolusjonsteori er tilstrekkelig til å forklare oppkomsten av nye funksjoner. Dette gjelder også for det andre eksemplet Coyne tar opp: rudimentære øyne hos huleboende dyr som fisk.
Det første eksemplet på et rudimentært trekk som er sitert her, er vingene til strutser og andre fugler som ikke kan fly, som strutsfugler. Få vil tvile på at en struts' forfedre var flygende fugler. Dette er imidlertid ikke et veldig sterkt bevis, hvis Coyne ønsker å overbevise skeptikere om effektiviteten til moderne evolusjonsteori - rudimentære strukturer av denne typen dokumenterer i beste fall tap av funksjon. Det som må demonstreres er at moderne evolusjonsteori er tilstrekkelig til å forklare oppkomsten av nye funksjoner. Dette gjelder også for det andre eksemplet Coyne tar opp: rudimentære øyne hos huleboende dyr som fisk.
Bilde 2. Lær av historiske feil -unngå junk-DNA feilen
Arrector pili, som får håret vårt til å reise seg, og som også refereres til av Coyne, er et annet eksempel på tap av funksjon (forfedrene våre var hårete enn oss). Uansett, selv hos mennesker, fungerer gåsehud til å bevare varme og dermed holde oss varme.
Coynes siste eksempel er musklene for å vrikke på ørene. Dette eksemplet representerer enda et tap av funksjon, snarere enn en gevinst. Kanskje våre forfedre brukte disse musklene til å bevege ørene og forbedre sjansene sine for å oppdage rovdyr. Men det betyr ikke nødvendigvis at vi deler en avstamning med andre nålevende primatarter.
Coyne forutser den typiske responsen på argumentet fra rudimentæritet:
"Motstandere av evolusjon reiser alltid det samme argumentet når 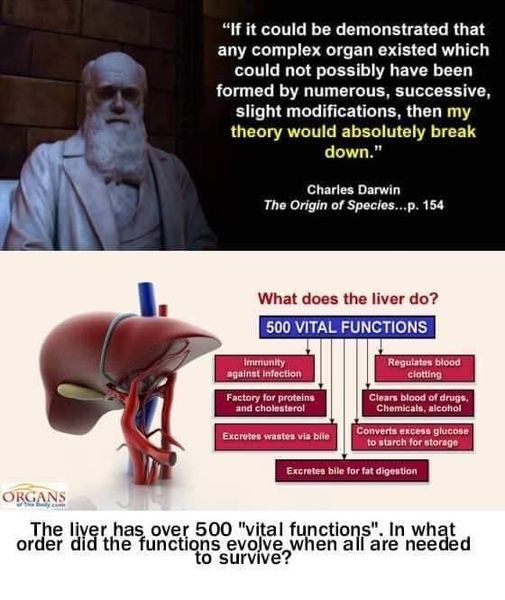 rudimentære trekk siteres som bevis for evolusjon. "Trekkene er ikke ubrukelige", sier de. "De er enten nyttige for noe, eller vi har ennå ikke oppdaget hva de er til for." De hevder med andre ord at et trekk ikke kan være rudimentært hvis det fortsatt har en funksjon, eller en funksjon som ennå ikke er funnet.
rudimentære trekk siteres som bevis for evolusjon. "Trekkene er ikke ubrukelige", sier de. "De er enten nyttige for noe, eller vi har ennå ikke oppdaget hva de er til for." De hevder med andre ord at et trekk ikke kan være rudimentært hvis det fortsatt har en funksjon, eller en funksjon som ennå ikke er funnet.
Men denne duplikken bommer på poenget. Evolusjonsteorien sier ikke at rudimentære karakterer ikke har noen funksjon. Et trekk kan være rudimentært og funksjonelt samtidig. Det er rudimentært ikke fordi det er funksjonsløst, men fordi det ikke lenger utfører funksjonen det utviklet seg for. (s. 58)"
Men etter Coynes beregning ville denne løse definisjonen av "rudimentæritet" sikkert innebære at ethvert organ og enhver struktur er rudimentær, siden alle trekk etter Coynes syn har utviklet seg fra noe annet. Som Jonathan Wells forklarer i sin egen anmeldelse av boken -lenke,
"Hvis menneskearmen utviklet seg fra beinet til et firfotet pattedyr (som darwinister hevder), så er menneskearmen rudimentær. Og hvis (som Coyne argumenterer) vingene til flygende fugler utviklet seg fra fjærkledde forbein til dinosaurer som brukte dem til andre formål, så er vingene til flygende fugler rudimentære. Dette er det motsatte av hva folk flest mener med "rudimentær".
Bilde 3. Motbevis mot trinnvis utvikling foreligger
Atavismer -lenke
Coynes neste emne er evolusjonære atavismer, som involverer det embryonale utseendet til et trekk som antas å ha vært tilstede hos en evolusjonær forfader. Selv om Haeckels teori om rekapitulering nå antas å være falsk, i det minste som et allestedsnærværende prinsipp, antas rekapitulering i denne mer begrensede forstand fortsatt å være gyldig. Coyne forklarer,
"Sanne atavismer må rekapitulere et forfedres trekk, og på en ganske nøyaktig måte. De er ikke bare monstre. Et menneske født med et ekstra ben, for eksempel, er ikke en atavisme fordi ingen av våre forfedre hadde fem lemmer. (s. 64)"
Med andre ord, når en embryonal abnormalitet samsvarer med en egenskap hos en evolusjonær forfader, kan fenomenet tas som bevis på felles aner. Men når en embryonal abnormalitet ikke samsvarer med en egenskap hos en evolusjonær forfader, teller ikke fenomenet som en "ekte atavisme". Faktisk blir mennesker født med abnormaliteter hele tiden, inkludert et ekstra ribbein eller en ekstra finger. Men ingen tror at dette gir bevis for at våre forfedre en gang hadde et ekstra ribbein eller en ekstra finger. Den samme sirkelformuleringen finnes rikelig i evolusjonær tenkning. Når likhet kan forklares med felles aner, kan den klassifiseres som "homologi" og dermed tas som bevis for felles aner. Men når likhet ikke kan forklares med felles aner, klassifiseres den som "analogi" og er bevis for konvergent evolusjon.
Eksempler på evolusjonære atavismer som Coyne presenterer inkluderer hvaler "født med et bakbein som stikker ut utenfor kroppsveggen", moderne hester "født med ekstra tær" og fugler som kan bli påvirket til å "produsere tannlignende strukturer" på nebbet. Alle disse eksemplene, som jeg ikke har noe problem med, illustrerer igjen organismers evne til å kvitte seg med egenskaper, men har liten relevans for hvor effektive de neodarwinistiske mekanismene er til å skape fundamentalt nye strukturer og egenskaper.
Døde gener
 Coyne vender deretter oppmerksomheten mot pseudogener. Ifølge Coyne kommer evolusjonen med en prediksjon:
Coyne vender deretter oppmerksomheten mot pseudogener. Ifølge Coyne kommer evolusjonen med en prediksjon:
Vi forventer å finne, i genomene til mange arter, tause eller "døde" gener: gener som en gang var nyttige, men som ikke lenger er intakte eller uttrykt. I motsetning til dette predikerer ideen om at alle arter ble skapt fra bunnen av at ingen slike gener ville eksistere, siden det ikke ville være noen felles forfedre der disse genene var aktive. (s. 66-67)
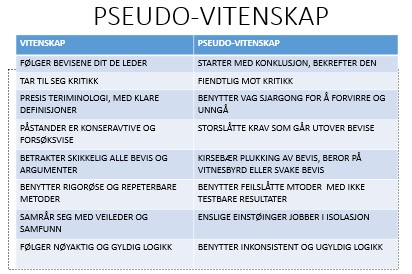
Bilde 4. Hvilken type vitenskap passer best med pseudo-vitenskap?
Han fortsrtter:
"Og den evolusjonære prediksjonen om at vi vil finne pseudogener har blitt oppfylt - i stor grad. Så å si alle arter har døde gener, mange av dem fortsatt aktive i sine slektninger. Dette innebærer at disse genene også var aktive i en felles forfader, og ble drept i noen etterkommere, men ikke i andre. Av omtrent 30 000 gener, for eksempel, bærer vi mennesker mer enn 2000 pseudogener. Vårt genom - og det til andre arter - er virkelig velbefolkede kirkegårder av døde gener. (s. 67)
Litteraturen det siste tiåret har imidlertid avdekket en mengde funksjoner for pseudogener. For å få en følelse av noen av funksjonene som er identifisert, ta en titt på følgende artikler:
"Pseudogener: Er de 'søppel' eller funksjonelt DNA?" (Balakirev og Ayala, 2003) -lenke
"Pseudogener: Pseudofunksjonelle eller nøkkelregulatorer innen helse og sykdom?" (Pink et al., 2011) -lenke
"Pseudogener er ikke lenger pseudo" (Wen et al., 2012) -lenke
"Pseudogener: Nylig oppdagede aktører i menneskelig kreft" (Policeno, 2012) -lenke
 Coynes fokus i denne delen er på det velkjente L-gulono-1,4-laktonoksidase (GULO)-genet, det siste enzymet i den biosyntetiske veien til askorbinsyre (vitamin C), og på luktreseptorgenene (OR) som er involvert i deteksjonen av luktmolekyler.
Coynes fokus i denne delen er på det velkjente L-gulono-1,4-laktonoksidase (GULO)-genet, det siste enzymet i den biosyntetiske veien til askorbinsyre (vitamin C), og på luktreseptorgenene (OR) som er involvert i deteksjonen av luktmolekyler.
Det fungerende GULO-genet lar de fleste planter og mange dyr produsere vitamin C fra glukose eller galaktose. I noen taxa fungerer imidlertid ikke GULO-genet i denne kapasiteten og får merkelappen "pseudogen". GULO-genet antas å være ødelagt hos mennesker (Nishikimi og Yagi, 1991), primater og marsvin (Nishikimi et al., 1994; Nishikimi et al., 1988), samt hos flaggermus av slekten Pteropus (Cui et al., 2011). Angående betydningen av GULO-pseudogenet med hensyn til felles aner, henviser jeg leserne til min forrige artikkel om emnet -lenke.
Bilde 5. GULO-gen gir opphav til spekulasjoner
Hva med menneskelige luktreseptor-pseudogener? Er de ikke-funksjonelle? Ikke nødvendigvis. Balakirev og Ayala (2003) bemerker faktisk at:
"De menneskelige luktreseptor (OR)-pseudogenene kan være viktige for generering og vedlikehold av reseptordiversitet. Intensiv intergenisk genkonvertering har blitt avslørt for denne multigenfamilien som fører til segmentomstokking i luktbindingsstedet, og en evolusjonær prosess som minner om somatisk kombinatorisk diversifisering i immunsystemet. Selv om OR-pseudogener har mistet sin fulle kodende funksjon, er de tilsynelatende underlagt nye evolusjonære begrensninger: OR-pseudogener tar i bruk ikke-kodende funksjoner som CpG-øyer, forsterkere og matriksfesteregioner."
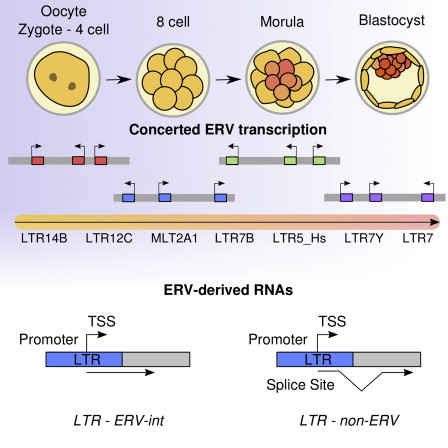 Coyne nevner også tilstedeværelsen av titusenvis av endogene retrovirus (ERV-er) i DNA-et vårt, som utgjør rundt 7 til 8 % av genomet vårt. Coyne avskriver disse viruslignende sekvensene som "døde gener". Forskningen det siste tiåret har imidlertid avdekket en rekke funksjoner for disse sekvensene. For et utvalg av noen av disse funksjonene, ta en titt på følgende artikler:
Coyne nevner også tilstedeværelsen av titusenvis av endogene retrovirus (ERV-er) i DNA-et vårt, som utgjør rundt 7 til 8 % av genomet vårt. Coyne avskriver disse viruslignende sekvensene som "døde gener". Forskningen det siste tiåret har imidlertid avdekket en rekke funksjoner for disse sekvensene. For et utvalg av noen av disse funksjonene, ta en titt på følgende artikler:
"Effekter av retrovirus på vertsgenomfunksjon" (Jern og Coffin, 2008) -lenke.
"Endogene retrovirus lange terminale repeats som bruksklare mobile promotorer: Tilfellet med primat β3GAL-T5" (Dunn et al., 2005) -lenke .
"Langtrekkende funksjon av et intergenisk retrotransposon" (Pi et al., 2010) -lenke .
"MuERV-L er et av de tidligst transkriberte genene i museembryoer som en gang var cellene" (Kigami et al., 2003) -lenke .
"Positivt utvalg av iris, et retroviralt konvoluttavledet vertsgen i Drosophila Melanogaster" (Malik og Henikoff, 2005) -lenke .
"Retrovirale promotorer i det menneskelige genomet" (Conley et al., 2008) -lenke .
"Artsspesifikke endogene retrovirus former transkripsjonsnettverket til det humane tumorsuppressorproteinet p53" (Wang et al., 2007) -lenke .
"Syncytin-A og syncytin-B, to fusogene placentaspesifikke murine konvoluttgener av retroviral opprinnelse konservert i Muridae" (Dupressoir et al., 2005) -lenke .
Coyne nevner at "noen av disse [retrovirale] restene sitter på nøyaktig samme sted på kromosomene til mennesker og sjimpanser." Jeg er imidlertid ikke overbevist om at det kan utelukkes at disse parallelle innsettingene kan gjenspeile preferensiell integrering av ERV-sekvenser. Slike målstedspreferanser har faktisk blitt identifisert for flere klasser av mobilt DNA (Spradling et al., 2011 -lenke ; Levy et al., 2009 -lenkek4/ , Li et al., 2009 -l79302 ; Zou and Voytas, 1997 C23835/ ; Wang et al., 2006 483006/ ).).
Bilde 6. RNA kan omformes via retrovirus
Palimpsester i embryoer
Coyne skriver:
"Lenge før Darwins tid var biologer opptatt med å studere både embryologi (hvordan et dyr utvikler seg) og komparativ anatomi (likheter og forskjeller i strukturen til forskjellige dyr). Arbeidet deres avdekket mange særegenheter som på den tiden ikke ga mening. For eksempel begynner alle virveldyr utviklingen på samme måte, og ser ganske ut som en embryonal fisk. Etter hvert som utviklingen går, begynner forskjellige arter å divergere - men på merkelige måter. Noen blodkar, nerver og organer som var tilstede i embryoene til alle arter i starten, forsvinner plutselig, mens andre går gjennom merkelige forstyrrelser og migrasjoner. Til slutt kulminerer utviklingsdansen i de svært forskjellige voksne formene for fisk, reptiler, fugler, amfibier og pattedyr. Likevel, når utviklingen begynner, ser de veldig like ut." (s. 73)
 Virveldyrembryoer, i motsetning til Coynes påstand, er faktisk vesentlig forskjellige i de tidlige stadiene. De konvergerer noe midtveis i utviklingen (på det fylotypiske stadiet eller "faryngula"-stadiet) før de divergerer igjen i senere utvikling. Som embryolog Brian Hall skriver i denne artikkelen: "Til tross for gjentatte påstander om ensartetheten til tidlige embryoer innenfor medlemmer av en rekke, er utviklingen før det fylotypiske stadiet svært variert." Faktisk viser selv mønstre av genuttrykk dette mønsteret (Kalinka et al., 2010 -lenke ).
Virveldyrembryoer, i motsetning til Coynes påstand, er faktisk vesentlig forskjellige i de tidlige stadiene. De konvergerer noe midtveis i utviklingen (på det fylotypiske stadiet eller "faryngula"-stadiet) før de divergerer igjen i senere utvikling. Som embryolog Brian Hall skriver i denne artikkelen: "Til tross for gjentatte påstander om ensartetheten til tidlige embryoer innenfor medlemmer av en rekke, er utviklingen før det fylotypiske stadiet svært variert." Faktisk viser selv mønstre av genuttrykk dette mønsteret (Kalinka et al., 2010 -lenke ).
Bilde 7. Embryoer i tidlig (ulik)-middels (likere)-sen(ulik) fase
Divergensen i virveldyrs embryonale utvikling i de tidligste stadiene er faktisk en grunnleggende utfordring for den konvensjonelle evolusjonære fortellingen. Embryonale screeningstudier viser at modifikasjoner i tidlig utvikling ikke tolereres av embryoer - konsekvensene er ofte dødelige (Kodoyianni et al., 1992 -lenke ).
Dessuten vil mutasjoner som påvirker kimcelleutviklingen sannsynligvis føre til infertilitet, en evolusjonær blindvei (Pellas et al., 1991). Likevel er embryoer ofte betydelig forskjellige med hensyn til dannelse av primordiale kimceller (PGC). Som Johnson et al. (2003) forklarer:
"Overraskende nok, blant de viktigste nålevende amfibielinjene, finnes én mekanisme [for PGC-dannelse] i urodeler [frosker og padder] og den andre i anuraner [salamandere og salamandere]. Hos anuranamfibier er PGC-er forhåndsbestemt av kimplasma; hos urodelamfibier dannes PGC-er ved å indusere signaler."
De bemerker videre:
"I anuranamfibier er primordiale kimceller (PGC-er), cellene som gir opphav til gameter, av endodermal opprinnelse, og de er spesifisert av den differensielle fordelingen av maternelt avsatte kimcelledeterminanter (kjent som kimplasma) til de antatte kimlinjeblastomerene. Dermed anses anuranamfibier å være forhåndsbestemt av kimplasma fra begynnelsen av utviklingen. Hos urodelembryoer stammer PGC-er fra lateral plate mesoderm. Urodele-embryoer inneholder ikke kimplasma, og derfor spesifiseres PGC-er senere i utviklingen enn hos anuraner. Hos urogenital deler dannes PGC-er som respons på ekstracellulære induserende signaler, ikke ulikt de som produserer andre mesodermale celletyper. Dette regnes som regulatorisk kimcellespesifikasjon."
Nå, her er gåten. Forskjellen er relatert til organer av ekstrem betydning - dvs. kimcellene. Forskjellen er ikke bare betydelig, men den forekommer ekstremt tidlig i utviklingen. Fra et evolusjonært rasjonale synspunkt ser det ut til å være to mulige forklaringer på dette:
En svært radikal endring har skjedd i de relevante utviklingsmekanismene.
En tidlig bifurkasjon -lenke har skjedd i fylogenien til amfibier - et bemerkelsesverdig tilfelle av konvergent evolusjon, kanskje fra forskjellige grupper av fisk.
Den andre hypotesen virker usannsynlig, for det ville være et bemerkelsesverdig sammentreff at to grupper hadde utviklet seg uavhengig over så lang tid og likevel delte så mange særegne trekk. Den første hypotesen virker enda mer usannsynlig siden påfølgende utvikling stammer fra disse tidlige prosessene. Som sådan vil modifisering av disse tidlige embryonale stadiene sannsynligvis forårsake katastrofal skade på organismen.
Dårlig design
 Argumentet om suboptimal design er beheftet med problemer. For det første, som nevnt tidligere, viser mange tilfeller av en gang antatt "dårlig design" seg ved nærmere ettersyn å vise designmessige avveininger. Dette gjør det til et argument fra uvitenhet, som må vike med hvert fremskritt i vår forståelse av disse systemene. Logikken er også usikker. Rettferdiggjør designfeilene i Windows-operativsystemet påstanden om at det ikke ble konstruert av intelligente agenter? Tydeligvis ikke. Evnen til å oppdage intelligensprodukter krever ikke at design er perfekt.
Argumentet om suboptimal design er beheftet med problemer. For det første, som nevnt tidligere, viser mange tilfeller av en gang antatt "dårlig design" seg ved nærmere ettersyn å vise designmessige avveininger. Dette gjør det til et argument fra uvitenhet, som må vike med hvert fremskritt i vår forståelse av disse systemene. Logikken er også usikker. Rettferdiggjør designfeilene i Windows-operativsystemet påstanden om at det ikke ble konstruert av intelligente agenter? Tydeligvis ikke. Evnen til å oppdage intelligensprodukter krever ikke at design er perfekt.
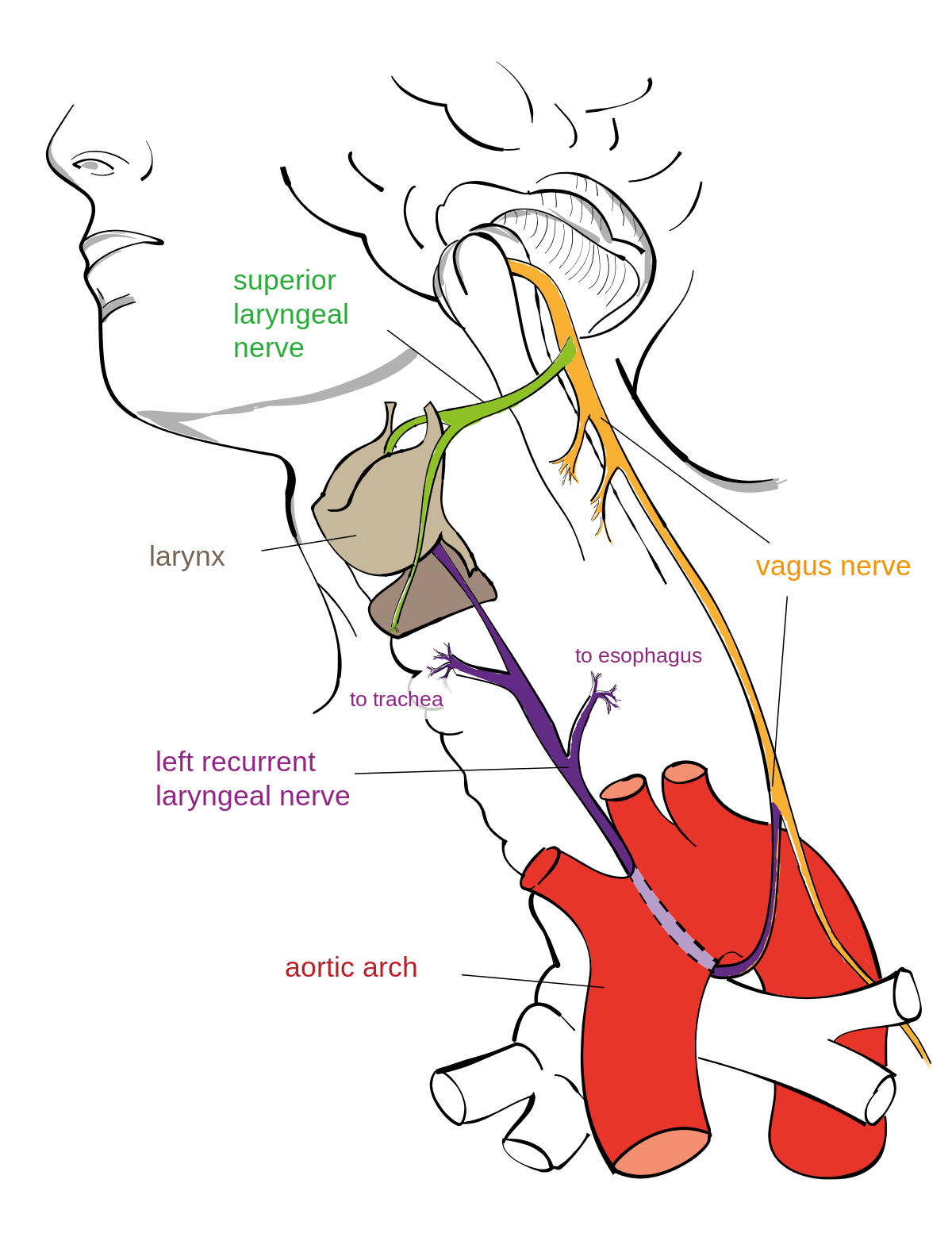
Coyne anser "en av naturens verste design" for å være den tilbakevendende laryngeale nerven (RLN), som sløyfer rundt aorta før den reiser tilbake opp til strupehodet i nakken. Hos sjiraffer ender denne omveien opp med å være 15 fot lengre enn den direkte ruten. Coyne forklarer denne tilsynelatende omveien med tanke på vår evolusjon fra fiskeforfedre. Men kan det være gode grunner til den tilsynelatende omveien til RLN? ENVs Casey Luskin gir oss tre mulige designgrunner for den valgte ruten:
Bilde 8. Hva slags vitenskap eller tro står vi for?
 1. Det finnes bevis for at antatte grunnleggende evolusjonære begrensninger som ville forhindret tap av den sirkulære ruten til RLN ikke eksisterer. Dette impliserer at det finnes en gunstig funksjon for den sirkulære ruten.
1. Det finnes bevis for at antatte grunnleggende evolusjonære begrensninger som ville forhindret tap av den sirkulære ruten til RLN ikke eksisterer. Dette impliserer at det finnes en gunstig funksjon for den sirkulære ruten.
2. RLNs bane lar den avgi filamenter til hjertet, til slimhinnene og til musklene i luftrøret på veien til strupehodet.
3. Det er dobbel innervasjon av strupehodet fra SLN og RLN, og faktisk innerverer SLN strupehodet direkte fra hjernen. Den direkte innervasjonen av strupehodet via den øvre laryngeale SLN viser at laryngeale innervasjoner faktisk følger selve designet som kreves av ID-kritikere som Jerry Coyne og Richard Dawkins. Ulike medisinske tilstander som oppstår når enten SLN eller RLN er skadet, peker på spesielle funksjoner for hver nerve, noe som indikerer at RLN har en spesifikk laryngefunksjon når alt fungerer som det skal. Denne segregeringen kan være nødvendig for å oppnå denne funksjonen, og redundansen ser ut til å bevare et visst nivå av funksjonalitet hvis én nerve blir skadet. Denne doble innervasjonen virker som et rasjonelt designprinsipp.
Bilde 9. Laryngeale nerve: Kan foreligge gode grunner for at den går som den gjør
Luskin siterer Michael Egnor, som lister opp noen andre interessante fordeler med ruten som RLN har tatt:
"Det er faktisk en designfordel med forløpet til de tilbakevendende nervene, hvis man ønsker å forfølge denne argumentasjonen. Nervenes forløp bringer dem gjennom mediastinum, der hjertet og lungene møtes. Det er mange lymfeknuter der, og forstørrelse av disse lymfeknutene fra prosesser som kreft eller infeksjon (f.eks. tuberkulose) irriterer ofte disse nervene og forårsaker heshet eller hoste. Nervenes forløp avslører sykdom i en ellers skjult del av kroppen (dypt inne i brystet) ved å forstyrre en prosess (tale) som er lett merkbar. Det tjener som en tidlig advarsel om å søke medisinsk behandling (eller, med smittsomme sykdommer, som en advarsel til andre om at denne personen er syk), og denne tidlige advarselen har reddet mange flere liv enn det redundante forløpet til nervene har kostet liv. Risiko/nytte-forholdet må undersøkes grundig før man hevder at nervenes forløp er biologisk ufordelaktig."
Egnor bemerker videre:
 "Selvfølgelig har ID-forkjempere aldri hevdet perfekt design. Men argumentet om at anatomien til den tilbakevendende larynxnerven er bevis på "dårlig design" feiler på mange nivåer. Nedstigningen av de tilbakevendende nervene under aortabuen og arteria subclavia er et resultat av koalesensmønstre og bevegelser av komponenter i aortabuen under embryogenesen. Det ser ut til at nærheten av forskjellige lag og strukturer i embryoet tjener til å veilede embryogenesen (det kalles induksjon). Detaljene i denne prosessen begynner bare å bli forstått, og det darwinistiske argumentet om at forholdet mellom de tilbakevendende nervene og aortabuen er bevis på dårlig design, tar ikke hensyn til den enorme kompleksiteten i embryonal utvikling. Det er analogt med en 3-åring som tar fra hverandre en datamaskin og hevder at den var dårlig designet fordi noen av kretskortmønstrene var "kurvede" i stedet for rette. Designvisdommen bak anatomien til de tilbakevendende nervene kan bare bedømmes av noen som kjenner alle designspesifikasjonene som er nødvendige for den delen av menneskekroppen. Selv de beste embryologene er førskolebarn når det gjelder det."
"Selvfølgelig har ID-forkjempere aldri hevdet perfekt design. Men argumentet om at anatomien til den tilbakevendende larynxnerven er bevis på "dårlig design" feiler på mange nivåer. Nedstigningen av de tilbakevendende nervene under aortabuen og arteria subclavia er et resultat av koalesensmønstre og bevegelser av komponenter i aortabuen under embryogenesen. Det ser ut til at nærheten av forskjellige lag og strukturer i embryoet tjener til å veilede embryogenesen (det kalles induksjon). Detaljene i denne prosessen begynner bare å bli forstått, og det darwinistiske argumentet om at forholdet mellom de tilbakevendende nervene og aortabuen er bevis på dårlig design, tar ikke hensyn til den enorme kompleksiteten i embryonal utvikling. Det er analogt med en 3-åring som tar fra hverandre en datamaskin og hevder at den var dårlig designet fordi noen av kretskortmønstrene var "kurvede" i stedet for rette. Designvisdommen bak anatomien til de tilbakevendende nervene kan bare bedømmes av noen som kjenner alle designspesifikasjonene som er nødvendige for den delen av menneskekroppen. Selv de beste embryologene er førskolebarn når det gjelder det."
 Bilde 10. Design-hypotese kan lede til videre forskning
Bilde 10. Design-hypotese kan lede til videre forskning
I et senere innlegg vil jeg vende meg til Coynes kapittel 4.
Jonathan McLatchie
Dr. Jonathan McLatchie har en bachelorgrad i rettsmedisinsk biologi fra University of Strathclyde, en mastergrad (M.RES) grad i evolusjonsbiologi fra University of Glasgow, en andre mastergrad i medisinsk og molekylær biovitenskap fra Newcastle University, og en doktorgrad i medisinsk og molekylær biovitenskap fra Newcastle University, og en doktorgrad i medisinsk biovitenskapelig biovitenskap fra University of Glasgow, og en doktorgrad, en mastergrad i medisinsk og molekylær biovitenskap fra University of Glasgow. i evolusjonsbiologi fra Newcastle University. Tidligere var Jonathan adjunkt i biologi ved Sattler College i Boston, Massachusetts. Jonathan har blitt intervjuet på podcaster og radioprogrammer inkludert "Utrolig?" På fremste Christian Radio, og mange andre. Jonathan har talt internasjonalt i Europa, Nord -Amerika, Sør -Afrika og Asia, og fremmet bevis for design i naturen.
Oversettelse, via google oversetter, og bilder ved Asbjørn E. Lund